BBC News, Русская служба - Главная
Главное

Live, Сырский рассказал об отступлении под Очеретино, но ISW считает, что быстрого прорыва РФ там не будет
Последние новости, комментарии и видео о войне России против Украины.

В Тбилиси снова протестовали против закона об «иноагентах». Произошли столкновения с полицией
В Тбилиси на проходящей у здания парламента многотысячной демонстрации против законопроекта об «иноагентах» накалилась обстановка. Сюда подтянут спецназ и дополнительные силы полиции.

В Германии зарезаны двое военнослужащих ВСУ. По подозрению в их убийстве арестован гражданин России
Украинское Министерство иностранных дел подтвердило, что два гражданина Украины, которые в субботу погибли от ножевых ранений в Германии, были военнослужащими ВСУ, проходившими реабилитацию. По подозрению в их убийстве арестован гражданин России.

Кошку в США случайно отправили курьерской почтой. Хозяйке пришлось лететь за ней тысячу километров
Американская семья нашла свою кошку после того, как ее случайно отправили курьерской почтой из штата Юта в Калифорнию.

Правительство Португалии отказалось платить репарации за колониальное прошлое. К этому призывал президент страны
Правительство Португалии заявило, что не планирует платить репарации за колониальное прошлое и активное участие в работорговле. Дискуссия об истории разгорелась в Португалии после того, как президент страны Марселу Ребелу де Соуза предложил властям заплатить за преступления прошлого.

Автосалон будущего. Как Китай становится главной автомобильной державой планеты
Китай решительно настроен завалить Запад машинами будущего, а Россию — машинами прошлого. И у него получается — почему?

В России арестованы еще двое журналистов. Их обвиняют в работе с командой Алексея Навального
Российские власти задержали двух журналистов по подозрению в сотрудничестве с Фондом борьбы с коррупцией Алексея Навального (который российские власти объявили экстремистской организацией).

На саммите в Саудовской Аравии снова пытаются запустить переговорный процесс между Израилем и ХАМАС
Попытки добиться прекращения огня между Израилем и ХАМАС, которое могло бы отложить израильское наступление на Рафах на юге Газы, продолжаются. Госсекретарь США Энтони Блинкен в Саудовской Аравии встречается с арабскими лидерами. Параллельно с этим египетская делегация накануне прибыла в Израиль в попытке реанимировать зашедшие в тупик переговоры.

Туалетная бумага на стебле: может ли растение заменить традиционный рулон?
Специалист-травник из Кении говорит, что туалетную бумагу можно заменить листьями. Эколог из США добавляет, что традиционный рулон - это те же растения, но после промышленной переработки

Военизированные чаты и наборы для выживания. Как молодые южные корейцы готовятся к войне с Севером
Все больше молодых людей в Южной Корее готовятся к войне с Севером. Помимо воды и продуктов питания, в наборы для выживания они кладут карту и компас, чтобы быть готовым к тому, что отключат мобильную связь или перестанет работать общественный транспорт.

В Огайо афроамериканец погиб после задержания. На записи полицейских он кричал: «Я не могу дышать»
Полиция Огайо опубликовала видеозапись задержания чернокожего мужчины, который впоследствии умер в больнице. Судя по обнародованным кадрам, он несколько раз говорил, что не может дышать. Американские медиа указывают на сходство этого происшествия с гибелью Джорджа Флойда, которая привела к массовым протестам в США.

«Контекст» — рассылка, которая помогает понять новости

Подкасты




Не пропустите

«Реальный американец, реально из Техаса». История Рассела Бентли, которого убили в Донецке
Волонтер из США Рассел Бентли последние 10 лет прожил в самопровозглашенной ДНР, а неделю назад стало известно, что он убит в Донецке российскими военными. Русская служба Би-би-си собрала все, что известно об этой истории.

«Разрушили все». Как российская оккупация навсегда изменила Чернобыльскую зону
Как изменилась Чернобыльская Зона за время войны в Украине.

«Мы тебя выдаем России, только не плачь». Почему политэмигрантам опасно в Казахстане, Кыргызстане и Армении и кто помогает им уехать
Страны, куда можно въехать по российскому паспорту — Казахстан, Кыргызстан и Армения, — после начала войны в Украине стали центрами притяжения россиян, в отношении которых в России по политическим мотивам заведены уголовные дела. Эти страны сложно назвать безопасными для российских политэмигрантов из-за тесных связей с Россией и общей базы розыска.

«Круг насилия бесконечен». Защищают ли Женевские конвенции жертв современных войн?
Чуть ли не каждый день мы слышим, что кто-то нарушает Женевские конвенции. Но мы совсем не слышим, что кто-то понес ответственность за эти нарушения: ни судов, ни арестов — только сплошные выражения озабоченности. Не настало ли время задуматься о пересмотре этих конвенций, раз они не работают?

Звонил гадалке и переписывался со знакомой. Что делал экс-министр экономики Казахстана Бишимбаев, пока его супруга умирала в ресторане
В Астане продолжается суд над экс-министром экономики Куандыком Бишимбаевым, обвиняемом в истязании и убийстве своей супруги Салтанат Нукеновой.

Какой будет война в Украине и российское наступление летом 2024-го
Где ждать самых ожесточенных боев, и какая военная техника будет иметь решающее значение для каждой из сторон? Анализируем перспективы боевых действий и описываем общее развитие событий российско-украинской войны в ближайшие месяцы.

«Обязательный минимум» классической литературы: список книг от Габриэля Гарсии Маркеса
По случаю десятой годовщины со дня смерти Габриэля Гарсии Маркеса — единственного колумбийца, удостоенного Нобелевской премии по литературе — редактор испаноязычной службы Би-би-си Эрнандо Альварес делится списком классических произведений, составленных писателем по его просьбе, который он бережно хранит вот уже почти 30 лет.

События

Американские университеты сотрясают антиизраильские протесты. В Лос-Анджелесе отменена выпускная церемония, дошло до столкновений с полицией
Масштабные протесты против действий Израиля в Газе не стихают в кампусах американских университетов. Критики протестующих обвиняют тех в антисемитизме. Администрации университетов пытаются найти тонкую грань между свободой слова и экстремизмом.

Франция получила Олимпийский огонь на церемонии в Афинах: фотографии
Олимпийский комитет Греции официально передал Франции огонь Летней Олимпиады 2024 года, которая пройдет в Париже с 26 июля по 11 августа.

Король Карл возвращается к публичной деятельности. Его лечение от рака проходит успешно
Король Великобритании Карл III на следующей неделе возвращается к публичной деятельности, сообщил официальный представитель Букингемского дворца.

Вашингтон призвал Пекин не содействовать России в войне, Китай предостерег США от пересечения «красных линий»
Министр иностранных дел КНР Ван И на встрече с госсекретарем США Энтони Блинкеном в пятницу в Пекине предостерег Вашингтон от пересечения «красных линий» в отношении Китая. А глава КНР Си Цзиньпин призвал США быть «партнерами, а не соперниками».

Двух британцев обвиняют в помощи российской разведке и организации поджога связанного с Украиной склада в Лондоне
Двум британцам официально предъявлены обвинения в содействии российским спецслужбам в связи с поджогом в Лондоне склада, принадлежащего компании выходцев из Украины.
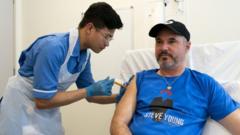
В Британии начались испытания первой в мире «индивидуальной» вакцины от меланомы
В Великобритании начались испытания первой в мире «индивидуальной» (то есть адаптированной под конкретного пациента) вакцины от меланомы — самой смертоносной формы рака кожи.

Эмма Стоун хочет, чтобы люди называли ее настоящим именем
При рождении девочку назвали Эмили, и именно это имя актриса считает по-настоящему своим. Но по всему миру ее знают как Эмму.

Лукашенко стал председателем ВНС; глава КГБ заявил, что по Минску хотели ударить дронами из Литвы. Главное в Беларуси за неделю
Русская служба Би-би-си рассказывает о главном в Беларуси за неделю

За граффити «Смерть режиму» школьницу из Читы приговорили к трем с половиной годам колонии
Первый Восточный окружной военный суд на выездном заседании приговорил читинскую десятиклассницу 17-летнюю Любовь Лизунову к трем с половиной годам колонии за граффити «Смерть режиму». Другие фигуранты дела Александр Снежков и Владислав Вишневский также осуждены.
Война России против Украины

США отправят Украине помощь на $6 млрд. Это крупнейший пакет за все время войны, но новых «Патриотов» в нем нет
Министерство обороны США заявило, что купит у военной промышленности военное оборудование для Украины на общую сумму 6 млрд долларов. Это крупнейшая одиночная партия помощи, которую Вашингтон направит Киеву за все время российского вторжения, и в него включены ракеты для американских систем ПВО Patriot. Однако Пентагон не сообщает о выделении самих пусковых установок Patriot, о которых в последнее время просила Украина.

Видео, «Все, что у меня есть, это мои руки, ноги и документы». Как новый закон о мобилизации затронул украинцев, живущих за границейПродолжительность, 4,31
23 апреля украинским мужчинам призывного возраста за рубежом временно перестали предоставлять консульские услуги. В паспортную службу Украины в Варшаве выстроилась очередь.

Разочарованные в «своем государстве». Что не так с ограничением прав украинских мужчин за границей
23 апреля украинским мужчинам призывного возраста за рубежом временно перестали предоставлять консульские услуги, хотя положения нового закона о мобилизации вступают в силу только 18 мая. Как мужчинам за рубежом обновить данные в ТЦК, обязательно ли для этого ехать в Украину? А как в таком случае пройти медкомиссию? Украинская служба Би-би-си пыталась прояснить эти вопросы с юристами.

Прорыв армии РФ у Очеретино в Донецкой области: чем это опасно для Украины
Пока украинские СМИ и соцсети выясняют, кто допустил беспрепятственный прорыв российских подразделений в село Очеретино к северо-западу от Авдеевки в Донецкой области, ситуация на этом участке фронта продолжает осложняться.

США тайно поставили Украине 300-километровые ракеты ATACMS. Их уже применили в Крыму
Соединенные Штаты в последние недели тайно поставили Украине ракеты 300-километрового радиуса действия ATACMS, и на прошлой неделе Украина впервые применила их в Крыму, сообщают американские чиновники и СМИ.

«Нас ждет тяжелый период, но Украина не проиграет». Интервью главы ГУР Кирилла Буданова Би-би-си
Кирилл Буданов — самый публичный руководитель военной разведки не только в истории Украины, но и в масштабах всего мира. Его прогнозы уже не такие оптимистичные, но, как утверждает он, Украина в войне, развязанной против нее Россией, не проиграет.

Не только через Тису. Украинец уплыл по Днепру на матраце в Беларусь, за что его осудили
В Беларуси судили украинца, незаконно переплывшего Днепр. Какими способами уклоняющиеся от мобилизации украинцы пытаются бежать из страны

«Это и есть Запорожская Сечь». Возродится ли Великий Луг на месте уничтоженной Каховской ГЭС
После уничтожения Каховской ГЭС, стоявшей на месте исторической части Запорожской сечи — Великого Луга, — многие опасались, что бывшее водохранилище превратится в пустыню или болото. Однако природа восстанавливается на удивление быстро, и властям предстоит решить, что с ней делать: восстанавливать ГЭС или превратить в заповедник.

Силовики пришли за продавцом шевронов для силовиков. Как волонтер и сотрудник парка «Патриот» стал обвиняемым в госизмене
Фигурантом нового дела о госизмене стал волонтер прокремлевского движения «Мы вместе», узнала Би-би-си. 26-летний Николай Мартынов во время войны с Украиной работал инженером в парке Минобороны «Патриот» и продавал шевроны с символикой силовиков. До ареста по «шпионской статье» суды пять раз подряд отправляли Мартынова в спецприемники — он провел там больше двух месяцев за якобы «нецензурную брань» и «неповиновение полиции».

Расследование Би-би-си: сколько может выжить на фронте в Украине российский заключенный?
За последний год заключенные, уехавшие на фронт из российских тюрем, принимали участие во всех ключевых боях на войне в Украине. Изучив истории более 9 тысяч отправленных на фронт осужденных, Би-би-си постаралась выяснить, как долго им удается выжить на войне и меняется ли этот показатель со временем.

«Такая двоякая ситуация». Как мать-одиночка из Сибири решила перевоспитать уголовника из ЧВК «Вагнер»
В России к обычной жизни стали возвращаться наемники ЧВК «Вагнер» — бывшие заключенные, помилованные Путиным в обмен на участие в войне в Украине, в том числе осужденные за тяжкие насильственные преступления. Би-би-си рассказывает, как жительница Красноярского края решила перевоспитать незнакомого ей ранее экс-наемника ЧВК «Вагнер».